
Cell Cycle
Cell Cycle and Mitosis
As we learned earlier, cells come from other cells. That means that a cell must grow, make a complete copy of its DNA, make more of everything else and finally divide that DNA exactly equally (and everything else fairly equally). This must be tightly regulated. In most cases, the two daughter cells created by cell division are identical (or, appear to be, at least).
Cell Cycle
We divide the cell cycle into stages based on what we could understand…and as we understand more, it gets more complex.
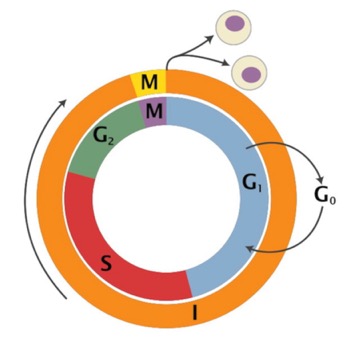
On this diagram, the outer circle is the oldest idea: There is mitosis and Cytokinesis (nuclear division and cell division, respectively) and then there is everything else (Interphase). We’ll talk about stages of mitosis in a bit.
Then, we recognized “S” phase, for DNA Synthesis, as another major event. The “gaps” where we couldn’t see much going on between Mitosis and S phase were called “G1” and “G2.” It turns out a lot of interesting regulation goes on there.
We also recognize something called “G0,” which means the cell is still alive, but no longer in the cell cycle…just sort of hanging out (that’s what most of the cells in your body are doing).
Here’s a look at the cycle from the standpoint of the DNA (Chromosomes). This odd little cell has a haploid number of chromosomes equal to 1 (the diploid has two, one from each parent, here shown as red or blue).
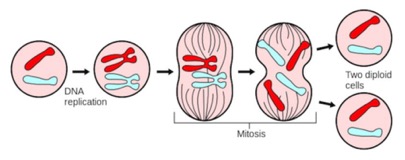
Normally, the DNA in the cell cannot be seen. Each chromosome is a long strand (each of your cells has a meter or so of DNA in its nucleus), but very narrow. Chromosomes are folded up into progressively higher orders. During mitosis, they become so tightly packed, you can see them in the cell.
Here is a great video. The only problem I have with it is it says at the end that chromosomes are not normally present in cells. I would argue they are present, but not in the tightly packed visible form (they would argue “chromosome” is the form you can see).
If you define "chromosome" as that thing you see at mitosis, I suppose that is right…they are only present during mitosis. That begs the question: what do you call the DNA when you cannot see it? I would call it a chromosome. We all understand “chromosome” refered originally to the mix of protein and DNA that is present in the mitotic cell. However, at this point, most modern biologists say chromosome when they mean the physical piece of DNA. The word “Chromatin” is often used to refer to the mixture of protein and DNA you cannot clearly see during interphase.
Checkpoints:
Coordination of all that has to go on requires that there are "checkpoints," where the cell assesses if it is safe to proceed (do I have enough nutrients to enter the very expensive "S" phase? Was DNA properly replicated so it is safe to go into mitosis?.
There are at least two “checkpoints,” where the cell must assess whether it is ready to go on. The “G1” checkpoint (called “start” in yeast) is the point where the cell commits to replicating its DNA. That’s an expensive proposition, so cells will only do that if they have the right signals. We talked about some of the types of signals a cell might need to grow and divide in the signaling section. If the signals (or food supply) are not there, the cells hangs out in G0. Once is gets the signal to pass this checkpoint, it’s a point of no return. It will try to finish DNA replication.
Since G1 is the period during which the cells undergo most growth. You could say the G1 checkpoint asks “Have I grown enough to enter S phase?”
How does it know if it has grown enough? Does it monitor total protein in the cell? Does it monitor cell mass or volume? Actually, it turns out it monitors the synthesis of one protein (or a couple) called cyclins. The presence of newly synthesized cyclins tell the cell each step is progressing well.
After DNA replication is completed, the cell has to assess if it is ready to divide. Maybe the DNA didn’t get replicated fully or is damaged. Maybe something has gone wrong with energy metabolism. If anything is wrong, the only other choice for the cell is apoptosis. We will talk about how p53 plays a big role in regulating this. So, this is the G2 checkpoint. If everything is fine, a signal is given and mitosis is initiated. Again, this is a “point of no return” for the cell. This is a “divide or die” decision.
The Signal:
Will it shock you to know that the two checkpoints are regulated by a kinase? In some organisms there are two or more different ones, in others, the same one is used for different checkpoints… and there are sometimes more than two checkpoints. The kinases are known as Cyclin-Dependent Kinases (or CDKs). A cyclin is the small protein made at a steady rate during different phases like G1 and G2. When the concentration of a cyclin gets high enough, it triggers activation of CDK proteins.
Analogous to G-proteins, cyclins bind to and activate the CDKs. However, there are some other proteins keeping the CDKs inactive. At some point, a “threshold” is reached, a “tipping point,” if you prefer, and enough Cyclin-CDK complex is around that the pathway becomes active and the cell passes the checkpoint. One of the effects of activating the Cyclin-CDK is this: it triggers the destruction of cyclin.
Does that remind you of anything? If cyclin is activating a target protein (in this case, the CDK) like the G-protein activates its targets, then the destruction of cyclin plays the same role as GTP hydrolysis. It turns off the signal so you can reset for the next cell cycle.
So, cyclin builds up…when enough of it is made, it activates the CDKs which results in passing the checkpoint AND the destruction of the cyclin. Cool, isn’t it.
It also won’t surprise you to know that several inhibitors of CDK are in clinical trials for treating cancer.
This link takes you to pages of links to structures of CDK2 and others in complex with dozens potential drugs designed to inhibit it. Should you want to, you can look at any of them with online viewers.
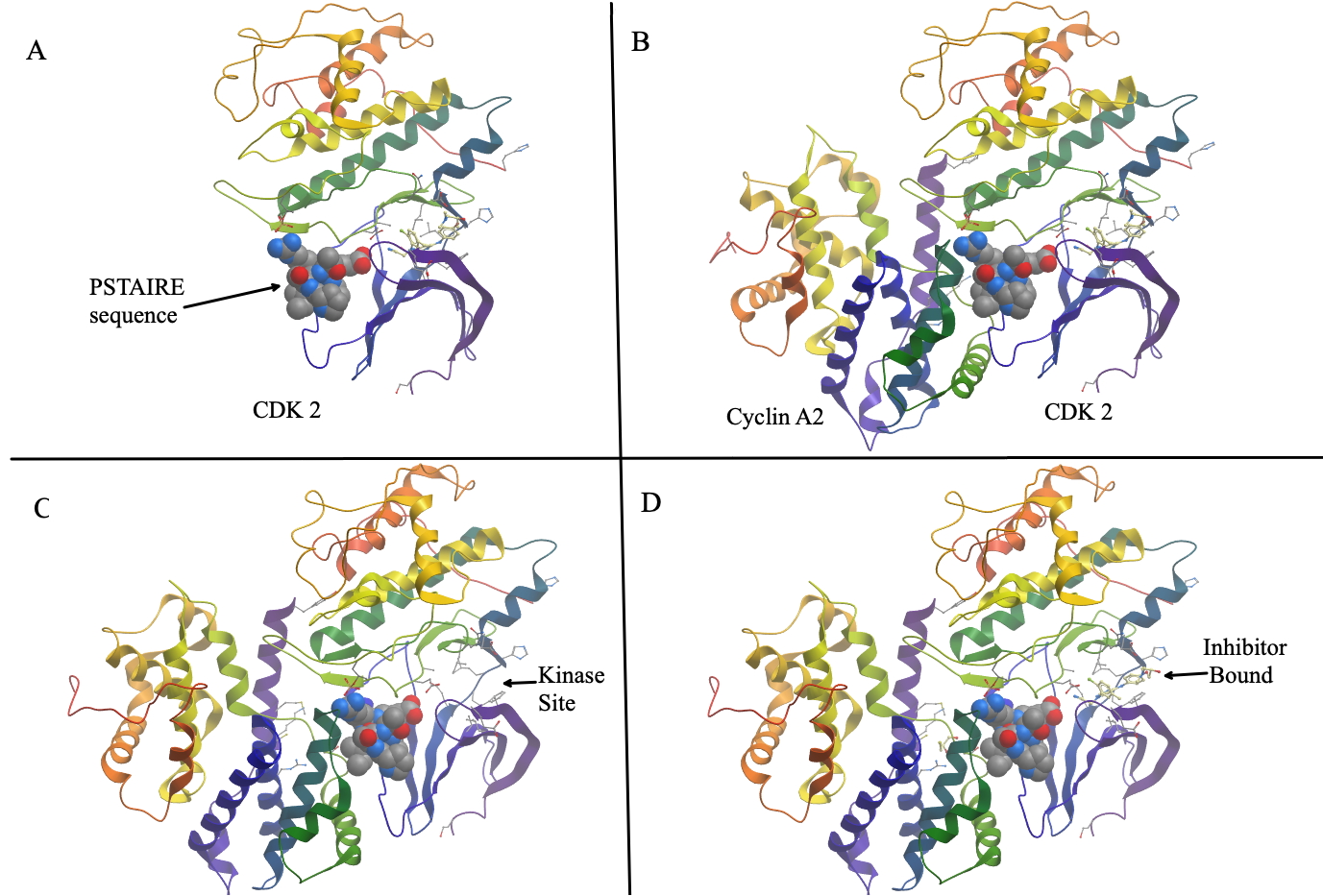
Below is a figure depicting the interaction of CDK2 with CyclinA2. Panel A shows a ribbon diagram of Cyclin A2. The alpha helix that mediates the interaction with cyclins is shown in space filling model. It is known as the PSTAIRE sequence. The name is just spelling out the single-letter code for the actual amino acid sequences: Pro-Ser-Thr-Ala-Ile-Arg-Glu- . Panel B shows the two proteins bound to each other. Figure is built from PDB entry 2iw6.
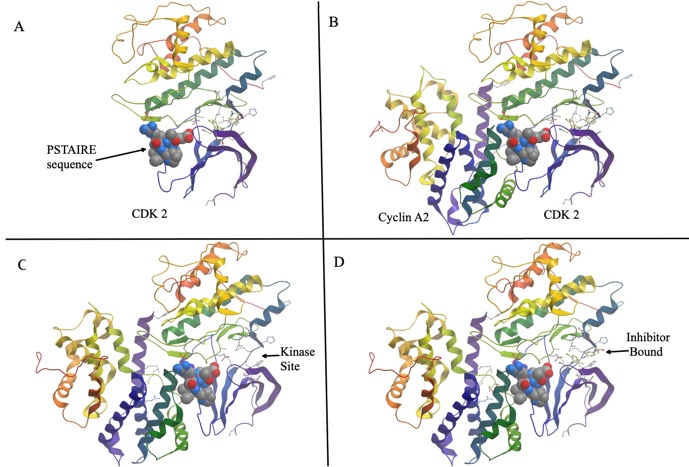 C and D highlight part of the active site without and with an inhibitory drug bound to it.
C and D highlight part of the active site without and with an inhibitory drug bound to it.The Frog Oocyte.
We talked in class about how Cyclin and CDKs were first seen as something called “Maturation Promoting Factor” (MPF), first isolated from frog oocytes. Here is the chart I went over today. It is similar to the one in the book on page 240. Note that in these cells, there is not much regulation of G1. The G1 checkpoint is sort of skipped because the frog oocytes don’t grow before dividing.
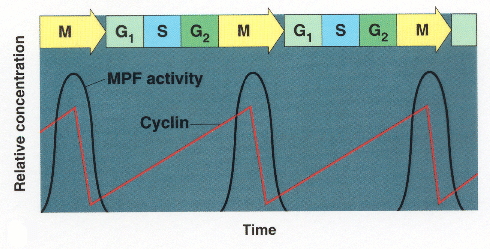
Note how the accumulation of cyclin is linear, more protein synthesized over time, but accumulation of MPF is not linear. The mechanism by which that threshold effect is interesting and we may get a chance to talk about it. But note that once MPF activity hits its peak, Cyclin is destroyed so that you can start over.
This is a reproduction of one of the first experiments published by Sir Tim Hunt. The dotted line is "cleavage index," a measure of the number of cells dividing. The line labeled "B" is the level of a non-cycling protein…I don't know what it is. It's just there to show that other proteins accumulate in a linear fashion with the growth of cells. The protein labeled "A" is a protein we now call "cyclin B."
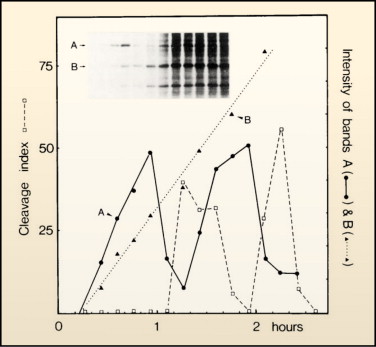
Note that the actual data show the peak of cycling several minutes before cells divide. But remember, the previous made-up chart was looking at amount of MPF in lysate, not when cells divide. After this checkpoint was worked out, it was relatively easy to find other cyclins. They all look similar at the gene sequence level and many were isolated as mutants in yeast. They work in a very similar fashion. There are several others that have been found. Each work with a CDK (sometimes the same kinase is used by several cyclins, sometimes there are unique Cyclin-dependent kinases). Here is a graph of intracellular concentration of various cyclins over the course of the cell cycle:
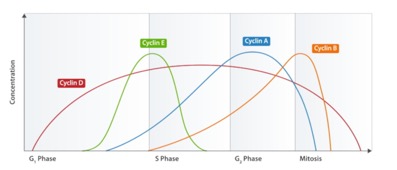
The one that was found in MPF is now known as Cyclin “B,” the mitotic cyclin. The others, in conjunction with CDKs, control other checkpoints. Cyclin “E” is the G1/S checkpoint cyclin, for example.
Stages of mitosis.
This isn’t hard, but you have to know them. I copied this from Indiana University’s biology site (go Hoosiers).
I like the diagram a lot. In “interphase, you cannot see the chromosomes. the really dark circles are actually nucleoli, where ribosomes are assembled. in prophase, you can see thick chromosomes forming. IN metaphase, they line up in the center. IN anaphase, they are being pulled to each “pole” (where the centrosomes are). Finally, Telophase, has the cell finishing division and the new nuclei forming.
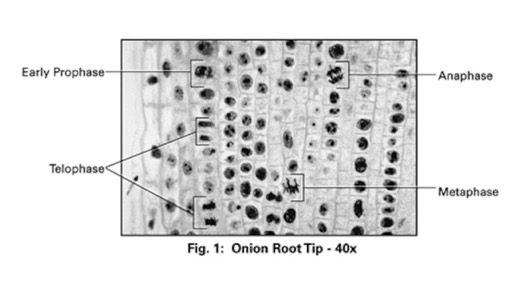
Through this process, each daughter cell gets a complete copy of the DNA. We’ll focus on the details of DNA replication later.
Here is a good video (embedded from youtube) that covers a lot of what we have discussed.
Meiosis
And here is a good one on Meiosis
The processes look fairly similar. In fact, meiosis II looks just like mitosis: Sister chromatids (really duplicated chromosomes) line up on the mitotic spindle and then separate. The different part is Meiosis I. Your cells are diploid (a complete set of chromosomes from Mom and another from Dad). Before you can pass your genetic information on to the next generation, cells of your germ-line (sperm or egg producing) must reduce the DNA precisely by half (and, of course, you have to finish college, get married and start a career etc).
It won’t do to just cut the amount of DNA in half. Each gamete must get one complete copy. Chromosomes are numbered from largest to smallest. So, chromosome 1 is the biggest. You have one of these from Mom and one from Dad. These are called homologous chromosomes. In Meiosis I, the duplicated homologous chromosomes pair up. So, both copies of chromosome 1, each really two copies of DNA as sister chromatids, will pair up, as will the two copies of chromosome 2 etc.
Meiosis 1 separates the mom copy of each from the dad. The assortment is random (each “mom” copy could go to either pole) The result will be a duplicated “haploid” cell, which has one duplicated copy of each chromosome. It could have mom’s chromosome 1 and Dad’s chromosome 2 or any combination.
In addition to this mixing up of mom’s and dad’s contribution, there is be some actual trading of DNA through a process called “crossing over” or “recombination,” which shuffles the genes a little during the time when homologous chromosomes are paired.
The videos show it pretty well.
